鲸,是地球上最大的哺乳动物。
它们巨大而可爱,智慧且优雅。
我们生活中经常能看到鲸豚的形象
时而它们是储蓄罐
时而是项链吊坠或耳饰
时而是一顶可爱的帽子
时而是某旅游类app的logo
面对如此可爱的动物,人们很难拒绝
这些熟悉又陌生的动物,你真的了解它们吗?
你知道,鲸在现生哺乳动物中最近的亲缘类群是什么动物吗?

根据分子*发生分析,鲸类与骆驼、猪、牛、羚羊等动物一样归入偶蹄目。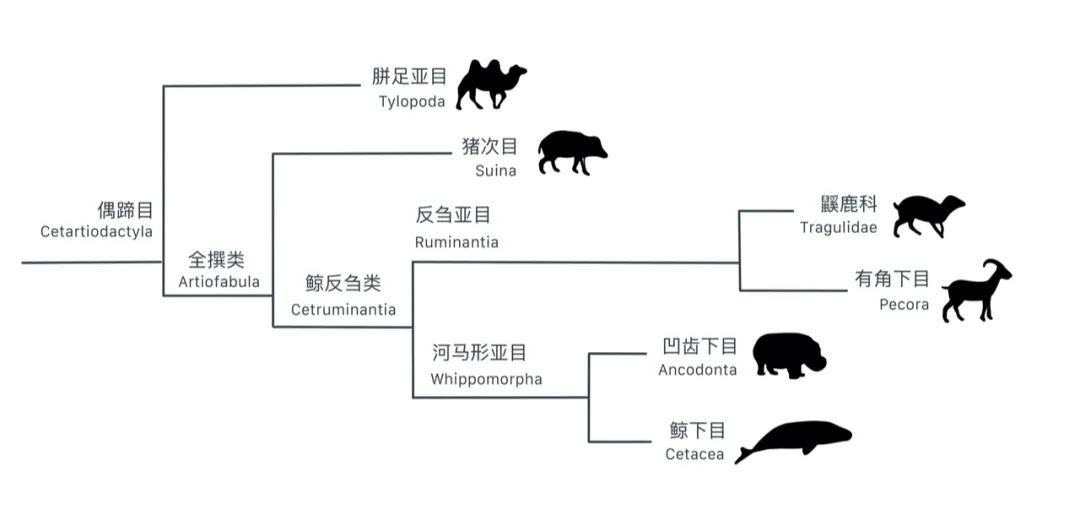
鲸的演化大致经历了六个阶段:
古偶蹄兽—巴基鲸—游走鲸—罗德侯鲸—龙王鲸—齿鲸与须鲸
鲸类是什么时候回到水中生活的呢?
鲸类的演化是为了开发一个空缺的生态位,这一生态位的产生可能是由于板块构造运动或5000 - 5500万年前发生的其他类型的环境变化。尽管当时陆生哺乳动物的多样性在不断增加,但海洋中几乎没有哺乳动物。
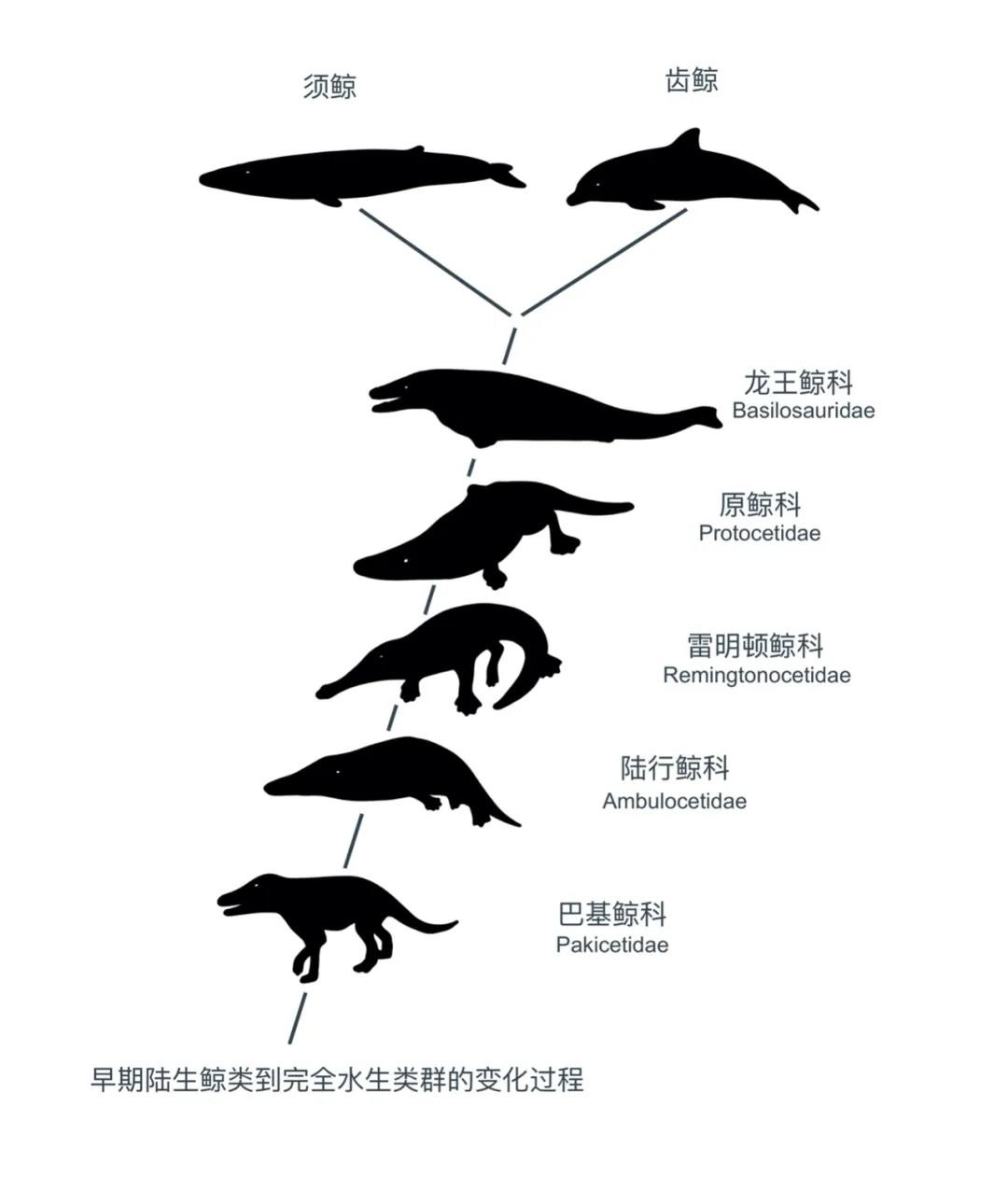
早期的鲸类及其近亲,如印原猪和巴基鲸都是陆生动物,大部分时间都在水畔活动。随着时间的推移,它们适应了海洋中的小生境。
走鲸和罗德侯鲸的化石显示出明显的游泳能力,尾巴扁平,后脚变大。另外,鼻孔从颅骨的末端逐渐移到头顶。
向水生栖息地的转移使这些物种得以开发陆地哺乳动物无法获得的资源,从而减少了对资源的竞争。竞争的减少使更多的个体得以生存和繁衍。
目前已知鲸类最早的姐妹群是4800万年前的印原猪,氧同位素δO18的数据及骨硬化性显示印原猪是在水中生活的,但δC13的数据却显示它们很少在水中觅食,这可能意味着鲸类最早的生活方式可能接近现存与鲸类最近亲缘的类群——河马:栖息于水中,但在水畔觅食。
随着时间的推进,鲸类在演化过程中越发趋于水生化,在目前已知最早的古鲸巴基鲸身上,眼的位置已经从侧面移动到更加靠近颅顶,利于从水中观察水面以上的环境,同时肢骨密度提升使得其能够更好地在浅水中活动,外耳痕迹也相应缩小。鲸的外耳最终消失,在水中的推进方式也由后肢推进转变为躯干——尾鳍推进,后肢和骨盆逐渐融合。等到始新世晚期的龙王鲸科身上,鲸类已经完全适应了水生生活。
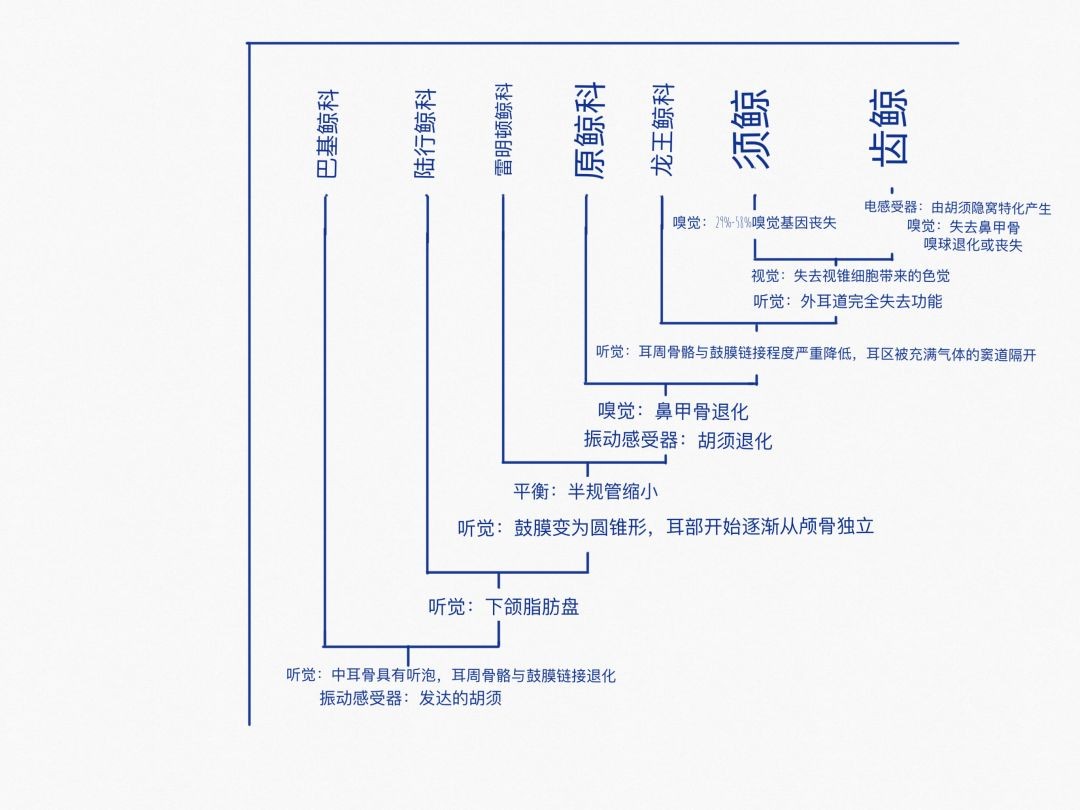
鲸类水生化过程中主要感官的变化
鲸类从陆地彻底重返水生环境仅仅花费了不到一千万年,这在漫长的地球生命史中仅仅是一瞬,但却对这一类群自身与全球的水域生态*产生了惊人的重塑作用。
代表性古鲸介绍
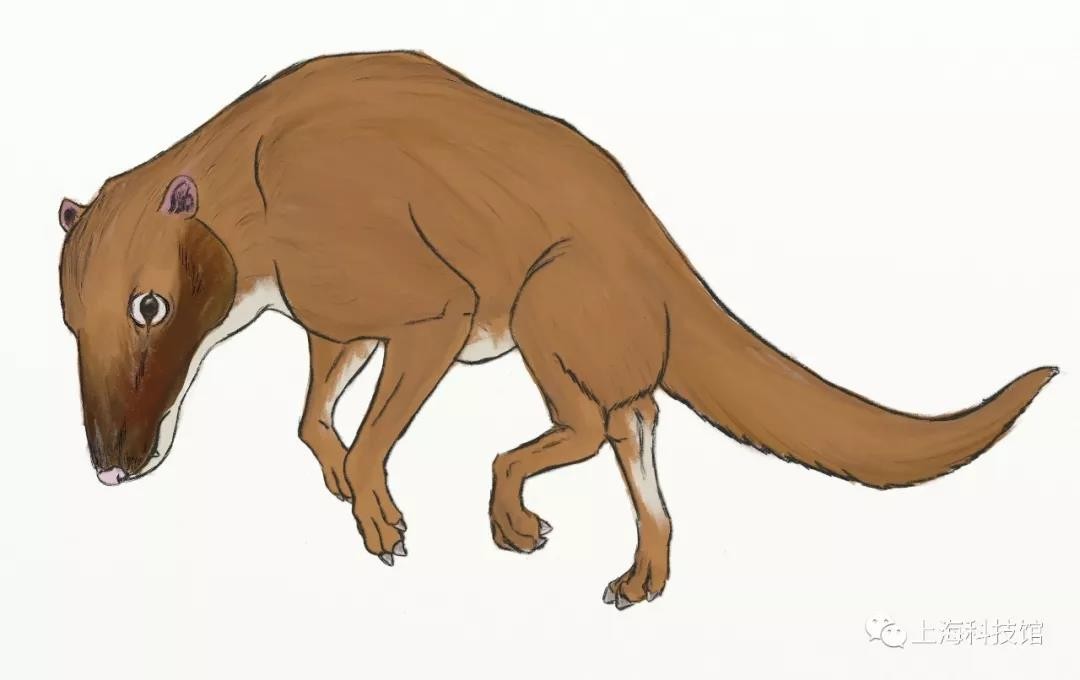
印原猪Indohyus
距今5000万年
化石发现于今天的印控克什米尔地区
体长约60厘米
印原猪复原图
图源:@秋山云
从体型和行为来说,印原猪接近现生的水鼷鹿Hyemoschus aquaticus。印原猪最重要的形态特征在于其耳骨具备中耳鼓泡,而这是鲸类演化支所独有的结构。同时通过对其骨骼密度以及体内氧同位素的分析,我们发现它实际上是一种半水生动物,其食物来源亦极可能包括底栖无脊椎动物。这一切化石证据说明了偶蹄目重返水中这一演化事件的发生时间实际上早于鲸类的产生,而空缺的生态位与额外的食物来源则成为了鲸类演化的重要动力。
巴基鲸Pakicetus
距今5200万年至4800万年
化石发现于今天的巴基斯坦
体长大约一到两米
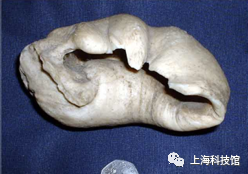
中耳骨照片
图源:维基百科
巴基鲸的化石所埋藏的地层形成于河流沉积物,这也说明了早期鲸类很可能首先进入的是淡水而非海洋环境。除此之外,巴基鲸的牙齿不再具备咀嚼面,这说明了其食性已经从杂食完全转变为了肉食性,而它的中耳鼓泡相比印原猪更加发达。在巴基鲸身上最重要的发现则是它的距骨具备偶蹄目所特有的双滑车构造。
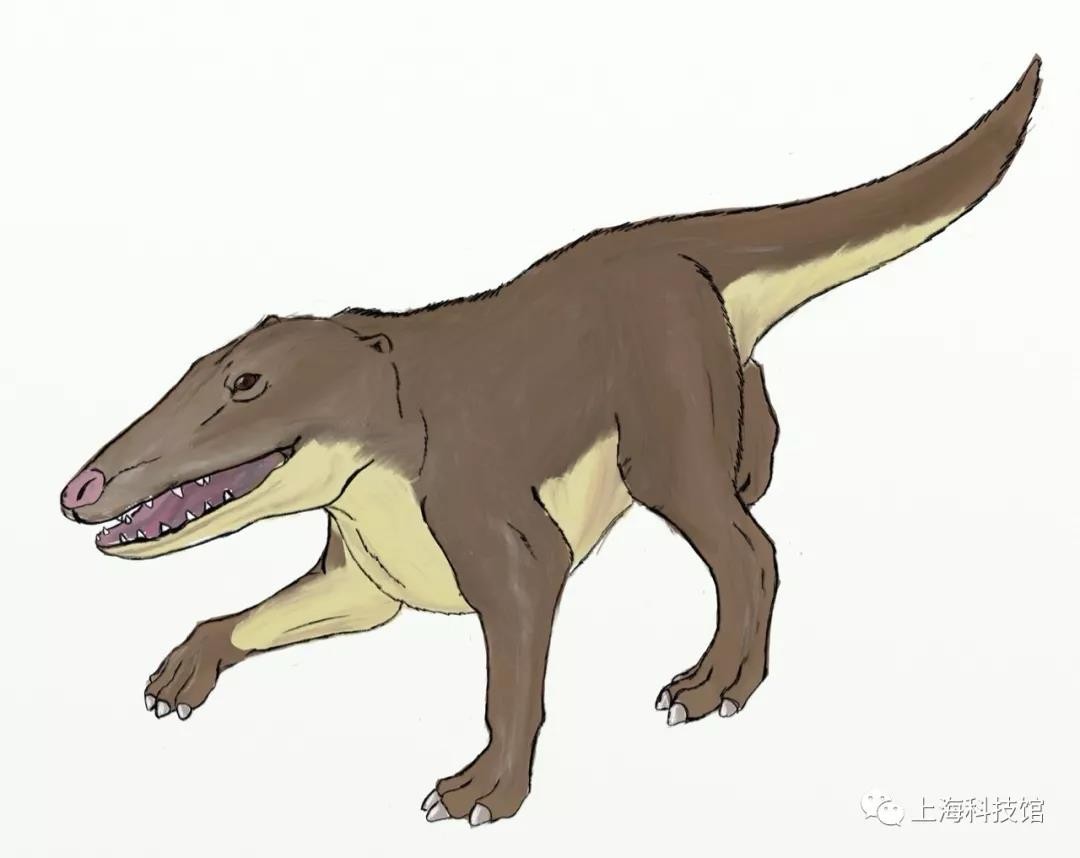
巴基鲸复原图
图源:@秋山云
走鲸Ambulocetus
距今4800~4000万年
化石发现于今天的巴基斯坦
体长约有3米

走鲸复原图
图源:@秋山云
走鲸科Ambulocetidae的模式种——游走鲸Ambulocetusnatans,它的化石完整度在已发现的古鲸小目化石中相当之高,其中最重要的部分则是该化石所保留的完整后肢。走鲸科的喜马拉雅鲸Himalayacetussubathuensis是我们已知最早进入海洋觅食的鲸类,但依然需要回到河流摄取淡水。
雷明顿鲸Remingtonocetus
距今4500-4350万年
化石发现于今天的印度与巴基斯坦
体长3.5米
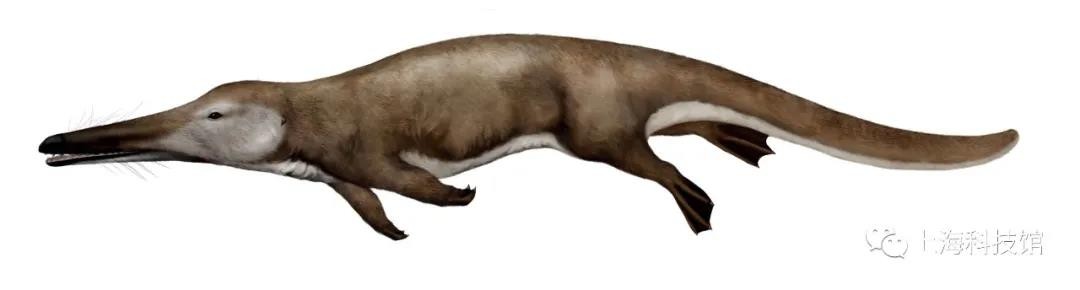
雷明顿鲸复原图
雷明顿鲸科是古鲸演化中的一个特殊旁支。雷明顿鲸最大的特征是它极其窄长的头骨,以比例而言,它有着比游走鲸更加短小的四肢和发达的尾部,因此科学家推测它水中的运动方式非常接近今天的巨水獭Castoroidesohioensis,更加依赖于尾与躯干的扭动。
慈母鲸Maiacetus
距今4700万年
化石发现于今天的巴基斯坦
体长约2.6米
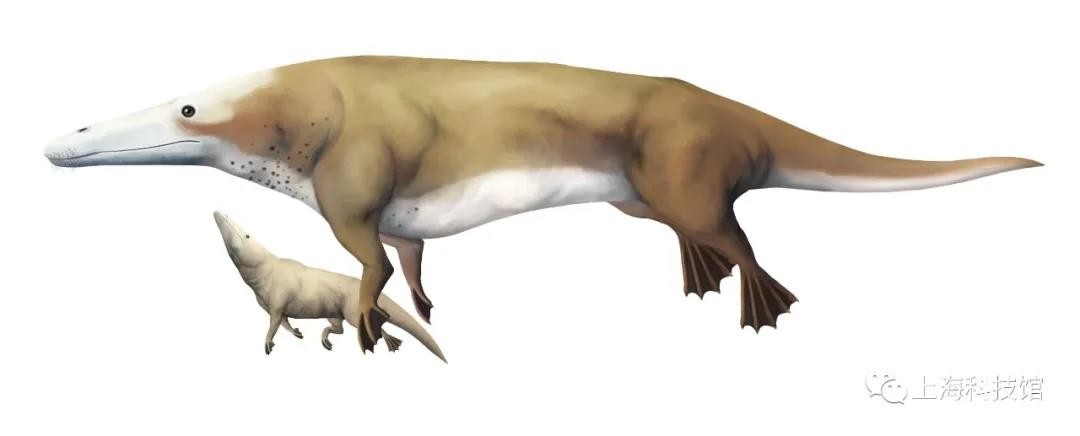
慈母鲸复原图
慈母鲸也是原鲸科的一员,共发现了一具完整的雄性骨架与保留有晚期胎儿的雌性骨架。这个物种提供了最早关于古鲸两性异型的化石证据(雄性体型更大且具备更加发达的犬齿),而雌鲸保留的发育晚期胎儿的头部朝向成鲸尾部,这与陆生偶蹄目相同却与现生鲸类相反,这也暗示了慈母鲸很可能依然在陆地上生育后代(因为水中生产时胎儿头部先离开产道会更加容易导致溺水)。
龙王鲸Basilosaus
距今4000万至3500万年
化石发现于今天的*斯安纳州
体长为15~18米
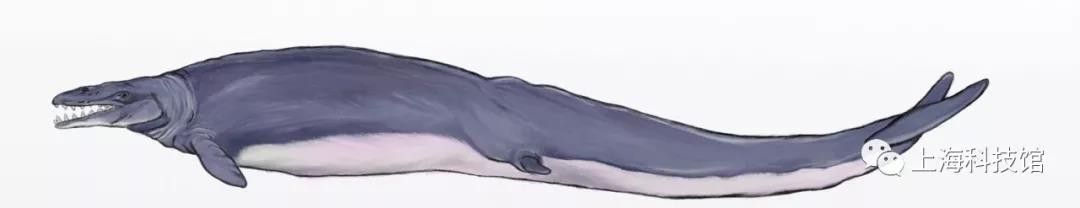
龙王鲸复原图
图源:@秋山云

鲸奇世界展览中1:1 龙王鲸复原图
图源:@秋山云
龙王鲸Basilosaus属于龙王鲸科Basilosaidae龙王鲸亚科Basilosa*inae,是该科的模式属,是人类最早发现的古鲸小目成员(发表于1834年),但由于发现之初被误认为是一种大型海洋爬行动物,不过著名的生物学家理查德·欧文于1839年将它正确地修订至鲸下目。龙王鲸是迄今为止已知体型最大的古鲸小目成员,它最鲜明的特征莫过于延长的躯干部分椎骨,这也使得龙王鲸的体型轮廓非常类似沧龙和鳗。龙王鲸也是古生物研究史中最早发现具有后肢和后足的鲸类,但它的后肢因比例大幅度缩小已经不再具有承重作用。
齿鲸和须鲸的分道扬镳
鲸类分为须鲸和齿鲸两类
齿鲸有圆锥形的牙齿,比如虎鲸。
须鲸只有鲸须没有尖锐的牙齿,比如蓝鲸。
大多数的学者认为须鲸与齿鲸是源自共同祖先,但是现在并没有确切的结论,通过多种研究科学家将分岐的时间定为2500万年前,分支的关键物种可能是矛齿鲸。
矛齿鲸Dorudon
距今3800万年~3400万年
化石发现于今天的北美洲、新西兰、撒哈拉西部与埃及的特提斯洋
体长约5米
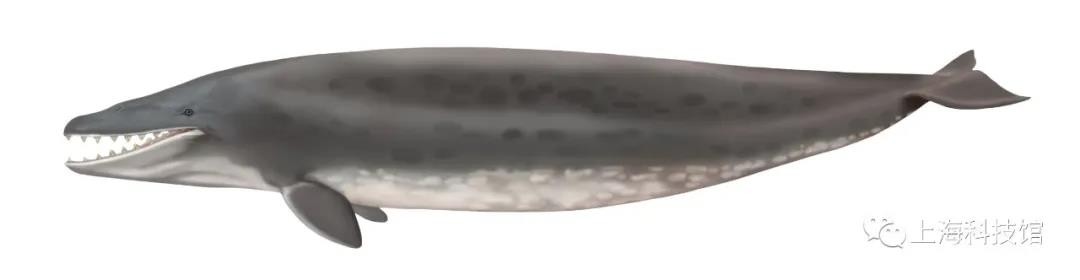
矛齿鲸复原图
矛齿鲸属Dorudon属于龙王鲸科下的矛齿鲸亚科Dorudontinae,模式种Dorudon serratus的模式标本发现于南卡罗莱纳州,包含了一个亚成体的部分头骨及躯干部分骨骼,之后陆续发现的多个个体的化石包含了这个物种全身的所有骨骼,这也使得这一类群成为了我们了解最为全面的古鲸之一。从*发生关系来说,矛齿鲸亚科中的某些物种极有可能是新鲸类(齿鲸+须鲸)的直系祖先,因而矛齿鲸属的化石其提供了关于现代鲸类祖先的生态学、分布、形态学甚至是软组织解剖学的大量信息。
须鲸演化的关键物种
拉诺鲸Llanocetus
距今3720万年~3390万年
化石发现于今天的南极洲西摩岛
体长8-10米

拉诺鲸复原图
图源:@秋山云
刻齿拉诺鲸Llanocetus denticrenatus是已知最古老的新鲸类之一,科学家于1989年在南极洲西摩岛上发现了它的化石,包括了部分下颌骨以及附在其上的两颗牙齿,拉氏鲸是人类已知第二古老的须鲸。刻齿鲸牙齿化石上发达的切割缘以及所有须鲸中最厚的牙釉质加上表面留下的摩擦痕迹说明它能够很好地切割而非单纯固定猎物。
绝大部分须鲸腭骨都具有腭沟,这一构造也被认为和鲸须的附着有关,然而拉氏鲸的这一构造却位于上牙槽骨之内,并为充分发育的牙龈提供血液,这一特征说明了腭沟不能作为鉴定是否存在鲸须的依据。
艾什欧鲸Aetiocetus
距今3300万年~2800万年
化石发现于*日本以及墨西哥
体长6米

艾什欧鲸复原图
视频源:@秋山云
艾什欧鲸属Aetiocetus,它是第一类被科学描述的带齿须鲸,填补了当时对于现代须鲸与古鲸祖先之间的认知空白,而2008年的研究表明它们同时具有牙齿与鲸须,这意味着艾什欧鲸科Aetiocetidae成为了鲸类演化重要的过渡类群。
大和鲸Yamatocetus
距今2810万年~2300万年
化石发现于今天的日本九州
体长7米

大和鲸复原图
图源:@秋山云
属于始须鲸科Eomysticetidae,大和鲸化石的特别之处在于完整度极高,其颌骨上保留了牙槽却没有任何牙齿,这也使得它成为了连接艾什欧鲸与无齿须鲸类Chaeomysticeti的重要纽带,它是通过一次含下大量海水再过滤方式捕食的,也是须鲸中已知最古老的物种。
齿鲸演化的关键物种
翘鼻鲸Simocetus
距今3390万年~2810万年
化石发现于今天的*俄勒冈州
体长约2米

翘鼻鲸复原图
图源:@秋山云
翘鼻鲸Simocetusrayi其属名词源自拉丁语Simus(意为形如八哥犬般短而翘起的鼻子),正是形容它所具有的短而向下弯曲的吻部,同时前颚骨上没有牙齿,这些构造可能是由于捕猎底栖猎物特化而成的。翘鼻鲸是已知齿鲸中最为古老的物种。
腔头鲸Cotylocara
距今2810万年~2303万年
化石发现于今天的*南卡罗莱纳州
体长约2米
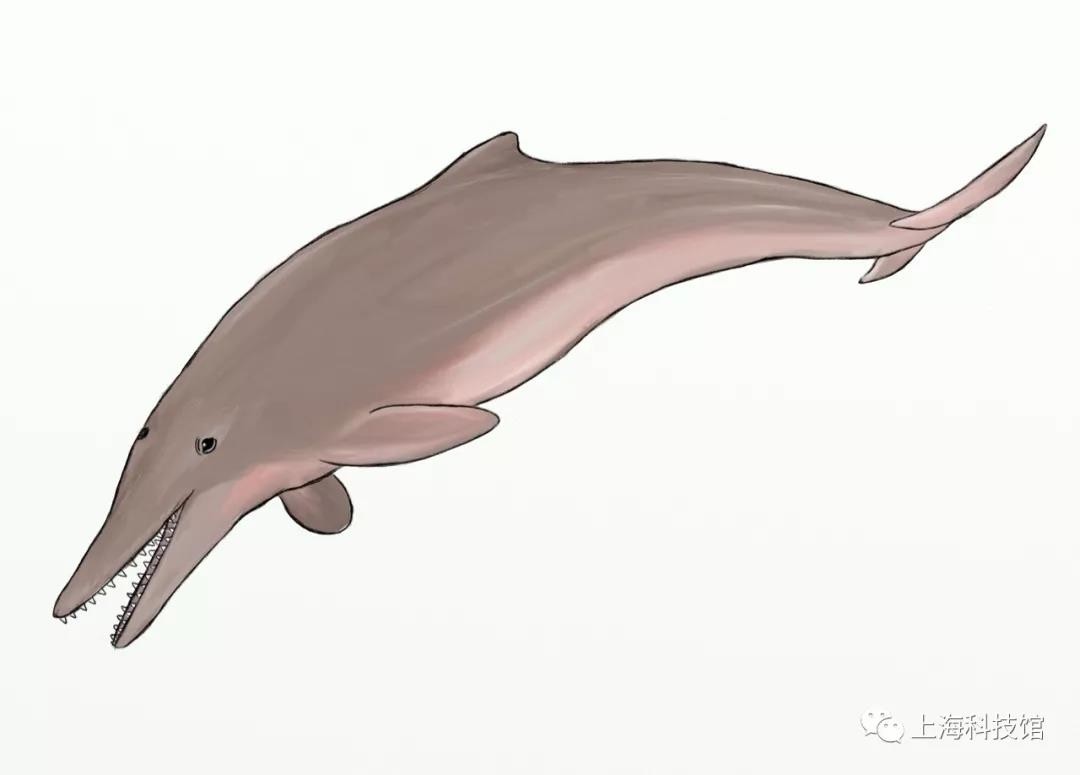
腔头鲸复原图
图源:@秋山云
其中代表性的梅氏腔头鲸Cotylocara macei的面部区域化石保留了关于早期齿鲸声纳演化的形态学证据,然而更为古老的翘鼻鲸和现代齿鲸的亲缘关系要近于腔头鲸,这意味着腔头鲸的声纳更可能是趋同演化的产物。
利维坦鲸Livyatan
梅氏利维坦鲸Livyatan melvillei
距今990万年~890万年
化石发现于秘鲁皮斯科伊卡
体长13.5-17.5米

利维坦鲸复原图
图源:@秋山云

鲸奇世界展览中的1:1利维坦鲸复原图及模型
图源:@秋山云
它的种名是为了纪念《白鲸记》的作者赫尔曼·梅尔维尔,它与作品的主角一样属于抹香鲸总科Physeteroidea。利维坦鲸的颅骨长度达到了惊人的3米,上下颚生长了大型的牙齿(部分牙齿的长度达到了36厘米),使它成为了最具有代表性的中新世掠食性抹香鲸之一。

鲸奇世界
《鲸奇世界》原创展览包括“大鱼传说”、“生存智慧”和“生态保护”三部分。分别从文化角度,呈现人类与鲸之间亘古以来的关联;从生物学角度,探索鲸的演化历程、生理习性与生存智慧;从生态学角度,关注鲸的生态保护,倡导公众保护海洋、江河,呵护水中精灵。

正在上海科技馆
地下一层临展厅
展出的《鲸奇世界》临展
让你了解更多鲸的故事
展览时间:1月18日至7月19日
展览地点:科技馆地下一层临展厅
参考文献
Waddell, P. J., Okada, N., & Hasegawa, M. (1999). Towards resolving the interordinal relationships of placental mammals. Systematic Biology, 48(1), 1-5.
Marx, F. G., Lambert, O., & Uhen, M. D. (2016). Cetacean paleobiology. John Wiley & Sons.
Marx, F. G., Lambert, O., & Uhen, M. D. (2016). Cetacean paleobiology. John Wiley & Sons.
Thewissen, J. G., Cooper, L. N., Clementz, M. T., Bajpai, S., & Tiwari, B. N. (2007). Whales originated from aquatic artiodactyls in the Eocene epoch of India. Nate, 450(7173), 1190.
Thewissen, J. G., Williams, E. M., Roe, L. J., & Hussain, S. T. (2001). Skeletons of terrestrial cetaceans and the relationship of whales to artiodactyls. Nate, 413(6853), 277.
Clementz, M. T., Goswami, A., Gingerich, P. D., & Koch, P. L. (2006). Isotopic records from early whales and sea cows: contrasting patterns of ecological transition. Jonal of Vertebrate Paleontology, 26(2), 355-370.
Thewissen, J. G., Cooper, L. N., Clementz, M. T., Bajpai, S., & Tiwari, B. N. (2007). Whales originated from aquatic artiodactyls in the Eocene epoch of India. Nate, 450(7173), 1190.
Thewissen, J. G., Cooper, L. N., Clementz, M. T., Bajpai, S., & Tiwari, B. N. (2007). Whales originated from aquatic artiodactyls in the Eocene epoch of India. Nate, 450(7173), 1190.
Gingerich, P. D., & Russell, D. E. (1981). Pakicetus inachus, a new archaeocete (Mammalia, Cetacea) from the early-middle Eocene Kuldana Formation of Kohat (Pakistan).
Thewissen, J. G., Williams, E. M., Roe, L. J., & Hussain, S. T. (2001). Skeletons of terrestrial cetaceans and the relationship of whales to artiodactyls. Nate, 413(6853), 277.
Thewissen, J. G., Hussain, S. T., & Arif, M. (1994). Fossil evidence for the origin of aquatic locomotion in archaeocete whales. Science, 263(5144), 210-212.
Bajpai, S., & Gingerich, P. D. (1998). A new Eocene archaeocete (Mammalia, Cetacea) from India and the time of origin of whales. Proceedings of the National Academy of Sciences, 95(26), 15464-15468.
Bebej, R. M., Zalmout, I. S., El-Aziz, A. A. A., Antar, M. S. M., & Gingerich, P. D. (2015). First remingtonocetid archaeocete (Mammalia, Cetacea) from the middle Eocene of Egypt with implications for biogeography and locomotion in early cetacean evolution. Jonal of Paleontology, 89(5), 882-893.
Gingerich, P. D. M. ul-Haq, WV Koenigswald, WJ Sanders, BH Smith, and IS Zalmout. 2009. New protocetid whale from the middle eocene of Pakistan: Birth on land, precocial development, and ual dimorphism. PLoS ONE, 4(2), e4366.
Owen, R. (1841). V.—Observations on the Basilosaus of Dr. Harlan (Zeuglodon cetoides, Owen). Transactions of the Geological Society of London, 2(1), 69-79.
Gingerich, P. D., Smith, B. H., & Simons, E. L. (1990). Hind limbs of Eocene Basilosaus: evidence of feet in whales. Science, 249(4965), 154-157.
Steeman, M. E., Hebsgaard, M. B., Fordyce, R. E., Ho, S. Y., Rabosky, D. L., Nielsen, R., ... & Willerslev, E. (2009). Radiation of extant cetaceans driven by restructing of the oceans. Systematic biology, 58(6), 573-585.
Milinkovitch, M. C. (1995). Molecular phylogeny of cetaceans prompts revision of morphological transformations. Trends in ecology & evolution, 10(8), 328-334.
Miller, D., & Jamieson, B. G. (2007). Reproductive Biology and Phylogeny of Cetacea: Whales, Porpoises and Dolphins.
Gibbes, R. W. (1845). Descript\\ion of the teeth of a new fossil animal found in the Green Sand of South Carolina.
Uhen, M. D. (2004). Form, function, and anatomy of Dorudon atrox (Mammalia, Cetacea): an archaeocete from the middle to late Eocene of Egypt.
Fordyce, R. E., & Barnes, L. G. (1994). The evolutionary history of whales and dolphins. Annual Review of Earth and Planetary Sciences, 22(1), 419-455.
Mitchell, E. D. (1989). A new cetacean from the late Eocene La Meseta formation Seymo Island, Antarctic peninsula. Canadian Jonal of Fisheries and Aquatic Sciences, 46(12), 2219-2235.
Fordyce, R. E., & Marx, F. G. (2018). Gigantism precedes filter feeding in baleen whale evolution. Crent Biology, 28(10), 1670-1676.
Emlong, D. (1966). A new archaic cetacean from the Oligocene of northwest Oregon.
Deméré, T. A., McGowen, M. R., Berta, A., & Gatesy, J. (2008). Morphological and molecular evidence for a stepwise evolutionary transition from teeth to baleen in mysticete whales. Systematic Biology, 57(1), 15-37.
Okazaki, Y. (2012). A new mysticete from the upper Oligocene Ashiya Group, Kyushu, Japan and its significance to mysticete evolution. Bulletin of the Kitakyushu Museum of Natal History and Human History Series A (Natal History), 10, 129-152.
Fordyce, R. E. (2002). Simocetus rayi (Odontoceti: Simocetidae, new family): A bizarre new archaic Oligocene dolphin from the eastern North Pacific. Smithsonian Contributions to Paleobiology, 93, 185-222.
Fordyce, R. E. (2008). "Neoceti". In Perrin, W. F.; Wsig, B.; Thewissen, J. G. M. (eds.). Encyclopedia of Marine Mammals (2 ed.). Academic Press. pp. 758–763. ISBN 978-0-12-373553-9.
Marx, F. G., Lambert, O., & Uhen, M. D. (2016). Cetacean paleobiology. John Wiley & Sons.
Lambert, O., Bianucci, G., Post, K., de Muizon, C., Salas-Gismondi, R., Urbina, M., & Reumer, J. (2010). The giant bite of a new raptorial sperm whale from the Miocene epoch of Peru. Nate, 466(7302), 105.